Brief description of bivalves. Class bivalve mollusks lamellar-branched. Bivalvia class - bivalvia
Mollusks are widespread secondary cavities, invertebrates. Their body is soft, undivided, in most it is divided into the head, trunk and leg. The main features of mollusks are the presence in most species lime shell and robes- a skin fold that covers the internal organs. The oral cavity of molluscs is filled with parenchyma. The circulatory system is not closed. Over 130,000 modern species and about the same number of fossil species are known. Mollusks are divided into classes: gastropods, bivalve, cephalopods.
class gastropods
class gastropods- this is the only class whose representatives have mastered not only water bodies, but also land, therefore, in terms of the number of mollusk species, this is the most numerous class. Its representatives are relatively small in size: Black Sea mollusk rapana up to 12 cm tall, grape snail- 8 cm, some naked slugs- up to 10 cm, large tropical species reach 60 cm.
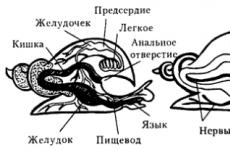
A typical class representative is big pond snail living in ponds, lakes, quiet backwaters. Its body is divided into a head, a torso, and a leg that occupies the entire ventral surface of the body (hence the name of the class).
The body of the mollusk is covered with a mantle and enclosed in a spirally twisted shell. The movement of the mollusk occurs due to the wave-like contraction of the leg muscle. A mouth is placed on the underside of the head, and on the sides are two sensitive tentacles, at their base are eyes.
The pond snail feeds on plant foods. In his throat there is a muscular tongue with numerous teeth on the underside, with which, like a grater, the pond snail scrapes off the soft tissues of plants. Through throat and esophagus food gets into stomach where it begins to digest. Further digestion takes place in liver and ends in the intestines. Undigested food is expelled through the anus to the outside.
The pond snail breathes with the help lung- a special pocket of the mantle, where air enters through the breathing hole. Since the pond snail breathes atmospheric air, it needs to rise to the surface of the water from time to time. The walls of the lung are braided with a net blood vessels. This is where the blood is enriched with oxygen and carbon dioxide is released.

Heart pond snail consists of two chambers - atrium and ventricle. Their walls alternately contract, pushing blood into the vessels. From large vessels through capillaries blood enters the space between the organs. This circulatory system is called open. From the body cavity, blood (venous - without oxygen) is collected in a vessel suitable for the lung, where it is enriched with oxygen, from where it enters the atrium, then into the ventricle and then along arteries- vessels carrying blood enriched with oxygen (arterial) enters the organs.
The excretory organ is bud. The blood flowing through it is freed from toxic metabolic products. From the kidney, these substances are excreted through the opening located next to the anal.
The nervous system is represented by five pairs ganglions located in different parts of the body, nerves depart from them to all organs.
Prudoviki are hermaphrodites, but their fertilization is cross. Eggs are laid on the surface of aquatic plants. They develop into juveniles. Development is direct.
The gastropods include slugs, named because of the abundant secretion of mucus. They don't have sinks. They live on land in humid places and feed on plants, fungi, some are found in vegetable gardens, causing harm to cultivated plants.
The herbivorous gastropods are grape snail also harmful to agriculture. In some countries it is used as food.

Among the numerous species of gastropods, sea shells are especially famous for their beautiful shells. They are used as souvenirs, buttons are made from the mother-of-pearl layer, and some peoples of Africa and Asia make money and jewelry from the shell of a very small kauri mollusk.
Bivalve class- exclusively aquatic animals. Through their mantle cavity, they pump water, choosing nutrients from it. This type of food is called filtration. It does not require special mobility of organisms, therefore, representatives of the class have some simplification in structure compared to representatives of other classes. All molluscs of this class have bivalve sink(hence the name of the class). The shell flaps are connected by a special elastic ligament located on the dorsal side of the mollusk. Muscles attached to shell valves contactors, their contraction contributes to the convergence of the valves, the closing of the shell, when they are relaxed, the shell opens.
Representatives of this class are , barley, oysters, mussels. The largest marine mollusk tridacna weighing up to 300 kg.

The most common mollusk in fresh water bodies of the country is. The body of a toothless, consisting of torso and legs, covered with a mantle hanging from the sides in the form of two folds.

Between the folds and the body there is a cavity in which gills and leg. Toothless has no head. At the posterior end of the body, both folds of the mantle are pressed against each other, forming two siphon: lower (input) and upper (output). Through the lower siphon, water enters the mantle cavity and washes the gills, which ensures breathing. With water, various protozoan unicellular algae, the remains of dead plants are brought. Filtered food particles pass through the mouth into stomach and intestines where they are exposed enzymes. The toothless is well developed liver whose ducts empty into the stomach.
Bivalves are used by humans. Mussels, oysters - are eaten, others, for example, are bred to obtain pearls and mother-of-pearl: pearl oyster, barley.
class cephalopods
Modern cephalopods There are about 700 species, exclusively inhabitants of the seas and oceans with a high concentration of salts, so they are not found either in the Black or in the Sea of Azov.
Cephalopods are medium to large sized predators. Their body is made up of torso and big head, the leg turned into tentacles that surround horn. Most of them have 8 identical tentacles, for example octopuses or 8 short and 2 long, like squid.
On the tentacles are suckers, with the help of which prey is retained. Only one tropical species does not have suckers - nautilus, but has a large number of tentacles. On the head of the representatives of the class there are large eyes resembling human eyes. Below, between the head and the body, there is a gap that connects with the mantle cavity. A special tube opens into this gap, called watering can, through which the mantle cavity is connected to the environment and is a modified part of the leg.

Many representatives of cephalopods do not have a shell, only the cuttlefish has it located under the skin, and the nautilus has a multi-chamber shell. The body is located in one of them, the others are filled with air, which contributes to the rapid buoyancy of animals. In many cephalopods, thanks to the jet mode of movement, the speed reaches 70 km per hour (squid).
The skin of many representatives of cephalopods is able to instantly change color under the influence of nerve impulses. Coloration can be protective (disguising itself as the color of the environment) or threatening (contrasting coloration, often changing). This is due to the high level of development nervous system, which has a complex brain, protected by a cartilaginous sheath - " scull”, sensory organs that determine complex behavior, in particular, the formation of conditioned reflexes.
For example, in case of danger, the salivary glands secrete poison that kills prey, or the ducts of the ink gland secrete a liquid that forms a black spot in the water; under its cover, the mollusk runs away from enemies.
Cephalopods are dioecious animals. They are characterized by direct development.
Cephalopods are of great industrial importance: they are used as food (squid, octopus, cuttlefish), brown paint is made from the contents of the ink bag of cuttlefish and squid - sepia, natural Chinese ink. In the intestines of sperm whales, a special substance is formed from the undigested remains of cephalopods - ambergris, which is used in the perfume industry to impart stability to the smell of perfume. Cephalopods are a food base for marine animals - pinnipeds, toothed whales, etc.
Tridacna. Pearls. Oysters. Scallops. mussels
Bivalves- marine and freshwater mollusks, which are characterized by the absence of a head, the presence of a wedge-shaped burrowing leg and the presence of a shell consisting of two wings. In attached species, the leg is reduced. Non-attached species can move slowly by extending their leg and then pulling their whole body towards it.
A mantle in the form of two folds of skin hangs down on the sides of the body of the mollusk. In the outer epithelium of the mantle there are glands that form shell valves. Substances in the valve are arranged in three layers: outer organic (conchiolin), calcareous and inner mother-of-pearl. On the dorsal side, the valves are connected by an elastic ligament (ligament) or lock. The sashes are closed with the help of closure muscles. On the dorsal side, the mantle grows together with the body of the mollusk. In some species, the free edges of the mantle grow together, forming holes - siphons for the input and output of water from the mantle cavity. The lower siphon is called the inlet, or gill, the upper one is the outlet, or cloacal.
In the mantle cavity on both sides of the leg are the respiratory organs - the gills. The inner surface of the mantle and the gills are covered with ciliated epithelium, the movement of the cilia of which creates a current of water. Through the lower siphon, water enters the mantle cavity, and out through the upper siphon.
According to the method of feeding, bivalves are filter feeders: food particles that have entered the mantle cavity are glued together and sent to the mouth opening of the mollusk, located at the base of the leg. Food from the mouth passes into the esophagus, which opens into the stomach. The midgut makes several bends at the base of the leg, then passes into the hindgut. The hindgut usually penetrates the ventricle of the heart and ends with the anus. The liver is large and surrounds the stomach on all sides. Bivalves, unlike gastropods, do not have a radula or salivary glands.
rice. one.
A - side view, B - transverse section: 1 - pedal ganglion, 2 - mouth,
3 - anterior muscle-contactor, 4 - cerebro-pleural ganglion,
5 - stomach, 6 - liver, 7 - anterior aorta, 8 - pericardium, 9 - heart,
10 - atrium, 11 - ventricle, 12 - posterior aorta, 13 - kidney,
14 - hindgut, 15 - posterior muscle-contact, 16 - viscero-
parietal ganglion, 17 - anus, 18 - mantle,
19 - gills, 20 - sex gland, 21 - midgut, 22 - leg,
23 - ligament, 24 - shell, 25 - mantle cavity.
The nervous system of bivalves is represented by three pairs of ganglia: 1) cerebro-pleural, 2) pedal, and 3) viscero-parietal ganglia. The cerebropleural ganglia are located near the esophagus, the pedal ganglia are in the leg, and the visceroparietal ganglia are under the posterior concha muscle. The sense organs are poorly developed. In the leg there are organs of balance - statocysts, at the base of the gills are osphradia (organs of chemical sense). Tactile receptors are scattered in the integument.
The circulatory system is an open type, consisting of the heart and blood vessels. The heart is three-chambered, has two atria and one ventricle. Blood from the ventricle enters the anterior and posterior aorta, which break up into small arteries, then the blood pours into the lacunae and is directed through the branchial vessels to the gills. Oxidized blood flows through the efferent gill vessels from each side of the body to its atrium and common ventricle.

rice. 2. bivalve larva
shellfish - veliger.
Excretory organs - two kidneys.
Bivalves are usually dioecious animals. The testicles and ovaries are paired. The genital ducts open into the mantle cavity. The spermatozoa are “ejected” by the males through the excretory siphon into the water and then are drawn through the introductory siphon into the mantle cavity of the females, where the eggs are fertilized.

rice. 3. Toothless larva
- glochidia:
1 - sashes, 2 - hooks,
3 - sticky (byssus).
In most species of bivalves, development occurs with metamorphosis. The planktonic larva veliger, or sailboat, develops from fertilized eggs (Fig. 2).

rice. four. Tridacna
(Tridacna gigas).
Giant Tridacna (Tridacna gigas)- the largest species of bivalves (Fig. 4). The mass of the tridacna reaches 250 kg, the body length is 1.5 m. It lives in the coral reefs of the Indian and Pacific oceans. Unlike other bivalves, the dorsal heavy part of the shell of the tridacna rests on the ground. This orientation of the shell led to great changes in the arrangement of various organs; in general, we can say that the tridacna turned 180 ° inside its shell. The only closing muscle has shifted to the ventral margin.
The edges of the mantle are greatly expanded and grow together almost throughout, except for three areas where the openings of two siphons and the opening for the exit of byssal filaments are located. In the thickened edge of the mantle live unicellular algae zooxanthellae. Tridacna is a filter feeder, but can also feed on these zooxanthellae.
The shells and meat of the tridacna have been used by the peoples of Oceania for a long time.
Pearls live in the Pacific and Indian Oceans at shallow depths (Fig. 5). They are fished for the purpose of obtaining pearls. The most valuable pearls are given by species of the genera Pinctada, Pteria.

rice. 5. pearl
(Pinctada sp.).
A pearl is formed if a foreign body (a grain of sand, a small animal, etc.) gets between the mantle and the inner surface of the mantle. The mantle begins to secrete mother-of-pearl, which coats this foreign body layer by layer, irritating her. The pearl increases in size, gradually separates from the inner surface of the shell and then lies freely. Often it does not connect to the sink from the very beginning. The pearl consists of alternating layers of mother-of-pearl and conchiolin. After 50-60 years after being extracted from the mollusk, it becomes covered with cracks, this is due to the destruction of the conchiolin layers inside it. The maximum period of "life" of a pearl as an ornament does not exceed 150 years.
In order to have a jewelry value, a pearl must have a certain size, shape, color, clarity. Pearls that meet "jewelry" requirements are rare in nature. At the end of the nineteenth century, a method was proposed for the artificial cultivation of pearls in sea pearls. Mother-of-pearl balls turned on a lathe are tied with sections of mantle sheets and transplanted into three-year-old mollusks in this form. The period of keeping the pearl sac ("nucleolus") is from 1 to 7 years.
At present, the technology of pearl breeding is as follows. Some farms grow pearl mussels up to the age of three, then transfer them to pearl farms. Here, pearl mussels are subjected to an operation (introduced "nucleoli") and then placed in special sieves, which are suspended from rafts. After a few years, the sieves are lifted and pearls are extracted from the pearls.

rice. 6. Oyster
(Crassostrea virginica).
Artificial breeding of marine animals is called mariculture.
oysters(Fig. 6) have been eaten by humans since time immemorial. The shell of oysters is uneven-valve: the left valve is larger in size than the right one and more convex. The left valve attaches the mollusk to the substrate. The mantle is open, does not form siphons, the flow of water is through. Well-developed semicircular gills surrounding a powerful adductor (muscle-terminator). Adult mollusks have no legs. Oysters are dioecious. Fertilized eggs develop in the posterior part of the mantle cavity of the female. After a few days, the larvae enter the water, swim, settle and attach themselves to the substrate. Oysters usually form clusters, distinguish between coastal settlements and oyster banks.
About 50 species of oysters are known, which belong to the families Ostreidae and Crassostreidae. One of the main commercial species is the edible oyster (Ostrea edulis). As a result of centuries of fishing, the number of oysters in many populations has declined sharply. Currently, along with fishing in natural habitats, oysters are grown artificially in specially organized oyster parks.
Oysters require specific conditions to grow. First, they feed on a certain type of plankton. Secondly, they do not live at a depth below 10 meters and at a water temperature below 5 ° C. Plantations are usually planted not very far from the coast in closed bays, so as not to be swept away by a storm. The growing period of oysters is not so short and is 34 years. Mollusks are kept in special containers, submerged to a certain depth and inaccessible to predators. After maturation, oysters are placed for a certain time in pools with clean sea water and special algae.

rice. 7.

rice. eight.
scallops- several dozen species of gastropod mollusks that belong to the families Pectinidae and Propeamusiidae. Scallops have a rounded shell with a straight locking edge, which has angled protrusions in the form of ears in front and behind. The surface of the valves has radial or concentric ribs. The leg is rudimentary, looks like a dense finger-like outgrowth. Numerous eyes and mantle tentacles with tactile receptors are located on the middle fold of the mantle (Fig. 7). Unlike other species of bivalves, scallops can swim by flapping their valves (Fig. 8). The slamming of the valves is provided by the contraction of the powerful adductor fibers. Scallops are dioecious animals.
The adductor of scallops, sometimes their mantle, is used for food. Just like oysters, scallops are not only hunted in their natural habitats, but also grown artificially (Patinopecten yessoensis). First, rafts are installed in the fenced area of the sea, to which collectors (pallets, panicles, etc.) are suspended. Mollusk larvae settle on these pallets. After 1-2 years, young mollusks are removed from the collectors, placed in individual nets and grown on "farms".

rice. 9. Mussel edible
(Mytilus edulis).
mussels- several species belonging to the Mytilidae family. They lead an attached lifestyle, in connection with which the leg is reduced, loses the ability to move and serves to isolate the byssal threads. The shell is of a characteristic "mytilid" shape, very dark in color, often blue-black. The shell of the edible mussel (Mytilus edulis) is about 7 cm long, up to 3.5 cm high, and 3.5 cm thick. The posterior adductor is much larger than the anterior one. Mussels are dioecious animals. Mussel settlements are a powerful biofilter that purifies and clarifies the water. It is estimated that mussels that settle on 1 m 2 of the bottom filter up to 280 m 3 of water per day.
Mussels are used for food. Fishing for these mollusks has been going on since ancient times. In addition, mussels are currently grown artificially. In this case, approximately the same technology is used as in the cultivation of scallops.
rice. ten. Teredo
(Teredo navalis):
1 - sink,
2 - body,
3 - siphons,
4 - moves, drilled
shellfish.
Teredo(Fig. 10) belongs to the family Woodworm (Teredinidae). The body shape is worm-like, so these mollusks have another name - shipworms. Body length up to 15 cm, at its anterior end there is a shell, reduced to two small plates. The sink is "equipped" with a drilling machine. At the posterior end of the body are long siphons. Hermaphrodites. In wooden underwater objects, the teredo "drills" numerous passages, feeds on wooden "crumbs". Digestion of wood is carried out by symbiotic bacteria. As a result of the activity of shipworms, the tree becomes like a sponge and is easily destroyed. Teredos pose a danger to wooden boats and buildings.
The class of bivalve mollusks, as is known, has four different names, each of which to some extent reflects the main features of their structure. Name "Bivalve"(Bivalvia) was first proposed by Linnaeus (1758) and is the most correct since it applies to all members of this class. Headless(Acephala), they were named by Link (1807), which reflected the fact that their head section of the body was reduced as they developed two shell valves in the process of evolution and these valves closed around the body of the mollusk. Third name - "lamellar"(Lamellibranchia), proposed by Blainville in 1814, can be fully applicable to only one detachment of this class, since the rest of the detachments have gills of a different structure; thus this title is inapplicable, as is the fourth - "axe-footed"(Pelecypoda, Goldfuss, 1880), since the structure of the leg in bivalve molluscs is very diverse. Thus, the most correct and comprehensive is the first, Linnean name, which should be retained in relation to this class.
Bivalves are widespread in the World Ocean and its marginal seas, in rivers and lakes, and even in ponds.
The total number of species of bivalves is about fifteen thousand, and most of them are associated in their habitat with salty sea waters, and only about one-fifth of the total number of their species inhabit fresh waters. On land, bivalves are not found.
In marine waters, they are extremely widespread, occurring in all climatic zones, from the warm waters of tropical seas to the Arctic and Antarctic and to the cold depths of the oceanic abyssal. They inhabit almost all the depths of the World Ocean - from the tidal zone (littoral) and coastal shallow waters up to the great depths of the depressions of the World Ocean, where they were: found at a depth of almost 10.8 km.
At present, the number of species of deep-sea bivalve mollusks living in the abyssal of the World Ocean (i.e., at a depth of more than 2000 m), judging by the still incomplete data, is about 400 species, but even this number should be considered greatly underestimated.
There is a great variety of sizes, structures and colors of shells of bivalve mollusks. So, a giant among mollusks in general, an inhabitant of tropical seas, a tridacna can reach 200 kg of weight, and the length of its powerful shell is 1.4 m. Along with this, the size of a number of ordinary deep-sea mollusks does not exceed 2-3 mm.
The shells and edges of the mantle of many tropical shallow-water mollusks, invisible among thickets of algae, white, pinkish, purple and yellowish corals, bright stars and other invertebrates, shine with colorful patterns and bright colors. A variety of outgrowths, spikes, scales and ribs adorn the shells of these mollusks, which helps them to strengthen themselves in these thickets and resist the action of sea waves and currents.
The shells of mollusks living on sandy or silty soils of the temperate or arctic region have a more modest color.
Deep-sea forms usually have a pale colored shell, often very thin and translucent.
Most of the freshwater forms are modestly colored in greenish and brownish tones.
The great diversity in the structure of the body and shell of bivalve molluscs is closely related to their way of life, their habitat, the depth and quality of the soil on which they live, or attach themselves, or burrow into. This affects primarily the structure of their shells; on the presence of ribs on it with one or another armament of its so-called lock, with the help of which the wings are fastened; on the presence or absence of siphons - special outgrowths of the mantle (two soft lobes that surround the body of the mollusk and secrete its shell); on the shape and size of the leg and the presence in it of a special gland that secretes the threads of the so-called b and c c c a with which they can be attached to the ground, as well as many other things. In forms that burrow more or less deeply into soft soil, special outgrowths of the mantle develop behind - siphons, through which water is sucked in and removed, which is necessary for breathing and feeding the mollusk immersed in the soil. These are the various makoms, tellins, yoldii and others. Mollusks living on the surface of the soil, crawling or slightly burrowing into it, have only rudimentary siphons or are completely devoid of them (for example, cockles, venusi, astartes and etc.). Mollusks that live in coastal shallow areas on harder sandy soils with an admixture of stones have stronger, thicker shells (for example, arches, scallops, sea scallops - pectenes and chlamys), and various inhabitants of soft silty soils have thinner shells ( batiarki, sea scallops propeamussiums and etc.).
Many forms living in coastal shallow waters attach byssus threads to stones, rocks, to each other, often forming whole clusters, intergrowths (many mussels), or even grow with their valves to stones, or grow together with each other ( oysters).
A very strong shell with sharp teeth on the ribs is possessed by many stone-boring clams; some of them secrete a special sour secretion that dissolves the lime of coastal rocks and stones in which they grind minks for themselves. Soft worm-like body of woodworm teredo(Teredo) is covered only in front by a small complex shell, which serves it for drilling, and not for protecting the body; spending their whole lives in tunnels gnawed through wood, these mollusks do not need to protect their weak, long body with a shell. Finally, a huge variety of various tropical mollusks, permanent inhabitants of coral reefs, is closely related to their life in a shallow, extremely diverse zone in terms of the nature of the substrate.
Strong calcareous shells of bivalves, as well as other mollusks, are well preserved in sediments (clays and sands) for entire geological epochs. The remains of their settlements are extremely valuable for geologists and paleontologists. These remains can perfectly characterize not only the hydrological and climatic conditions under which these deposits were formed (i.e., under which the species of mollusks found here lived), but also the age of the given sequence of deposits. Thus, accumulations of fossil shells of a cold-water mollusk now living in the Arctic seas portland arctic(Portlandia arctica) in the deposits of the north of Europe perfectly indicate that these areas were formerly occupied by the cold, slightly freshened waters of the shallow so-called Yoldiev Sea. This sea with a cold-water fauna, where the Arctic Portlandia played a leading role, was associated with a period of cooling in the post-glacial period (about 8-10 thousand years ago). Conversely, the deposits of the warm Littorina Sea, which formed later (3-5 thousand years ago), are characterized by the presence of remains of completely different, warm-water mollusks, such as Icelandic cyprina(Cyprina islandica), edible cockles(Cerastoderma edu1e), cirphei comb(Zirfaea crispata) and others. These species now live only in the North Atlantic, in the warmest regions of the Barents and partly the White Seas, while in the era of the Littorina Sea they moved further north.
Representatives of the class of bivalve mollusks first appear in deposits in the Paleozoic, that is, in the oldest deposits of our planet, namely in the Upper Cambrian layers, the formation of which dates back about 450-500 million years. The first bivalve mollusks found here belonged to four genera, of which such as Ctenodonta and Paleoneilo had a comb lock and outwardly resembled modern walnut(Nuculidae) and mallecium(Malletiidae) from comb-toothed order(Tachodonta). The greatest species diversity of bivalve molluscs reached in the Cretaceous, ie, 100-130 million years before our time.
Thus, the class of bivalves is one of the most ancient groups of benthic invertebrates.
Since ancient times, many bivalve molluscs have been used by humans, they have served and are still being prey. Their shells are constantly found in the so-called "kitchen heaps" of prehistoric man, who lived near the shores of the seas, rivers, lakes. In the excavations of Paleolithic human sites in the Crimea, a large number of shells of oysters, mussels, scallops and other mollusks, which are hunted to this day, are invariably found. Bivalve molluscs are harvested for their tasty, very healthy and easily digestible meat (such as oysters, mussels, scallops, tapes and venerupis cockerel, mactra, sand shell, cockles, arches, sea cuttings and synovacules, freshwater perlovitz, lampsilia, toothless, corbicula and etc.).
In terms of calories, they can even surpass the meat of many fish, both marine and freshwater. The nutritional value of shellfish meat is also determined by the high content of vitamins A, B, C, D, etc., high content such rare minerals in ordinary human food as iodine, iron, zinc, copper, etc. The latter, as you know, being part of a number of enzymes, hormones, play an extremely important role in oxidative, carbohydrate and protein metabolism, in the regulation of hormonal activity . The meat and shells of molluscs are widely used for the manufacture of fodder flour for fattening poultry, as well as for the manufacture of fertilizer fats.
In recent decades, due to the fact that the natural reserves of the most valuable edible mollusks (even in the seas) are depleted, and the demand for them continues to increase, in many countries they began to be relocated to new areas, acclimatized, and also bred artificially both in marine and and in fresh waters, on "farms" - specially prepared shallows and in small bays protected from predators, artificial reservoirs. Successfully bred and cultivated not only marine mollusks (oysters, mussels, barnacles, tapes), but also freshwater (lampsilin).
Currently, significantly more than half of the harvested bivalve mollusks are obtained as a result of their artificial breeding. The capture of mollusks in their natural habitats in water bodies and their artificial breeding have now become a profitable and important part of the food industry in a number of countries.
Bivalves are now often caught on large vessels with specially designed fishing gear; scuba diving is widely used. Shellfish enter the market not only in fresh and dried form, but especially in ice cream; the preparation of various canned molluscs also developed greatly.
The extraction of bivalve mollusks has increased dramatically in recent decades. If before the start of the World War their annual production was about 5 million cu, then already in 1962 it increased to about 17 million cu and began to account for about 50% of the world production of all marine invertebrates, or 4% of the total world production (426 million v) all products of the sea (fish, whales, invertebrates, algae).
The largest number (about 90%) of bivalve mollusks is mined in the northern hemisphere - in the Pacific and Atlantic oceans. Fishing for freshwater bivalve mollusks provides only a few percent of their total world production. Of particular importance is the bivalve fishery in countries such as Japan, the United States, Korea, China, Indonesia, the Philippine Islands and other Pacific islands. Thus, about 90 species of bivalve mollusks are mined in Japan, of which about two dozen species are of great commercial importance, and 10 species are artificially bred. In European countries, the fishing and breeding of bivalve mollusks is most developed in France, Italy, etc.
In the USSR, commercial importance is mainly large seaside scallop(Pecten (Patinopecten) yessoensis), as well as various mussels, white shell(Spisula sachalinensis), sand shell(Mua (Arenomya) arenaria), cocks(Tapes, Venerupis) and some others.
Bivalve mollusks have long been mined for the sake of their shells, which provide not only excellent raw materials for mother-of-pearl products (many freshwater pearls and pearl mussels, sea pearls - pinctadas, pteria, etc.) ”but also the most valuable pearls. At the beginning of this century, industrial methods were found to more quickly artificially obtain pearls (the finds of which are a rather rare accident under natural conditions), indistinguishable from naturally formed pearls. Farms for keeping sea pearls and growing pearls in them are especially successful in Japan. So, already in 1936, 140 thousand sea pearl shells were grown here and 26.5 thousand pearls were obtained.
In some countries, especially in the tropical Pacific, bivalve shells are widely used for lime production.
Almost all bivalve mollusks, with the exception of large forms with a strong, thick shell, serve as a favorite food for bottom fish - benthophages (i.e., feeding on bottom animals), including many commercial fish, both marine and freshwater: flounders, some cod (hadock), sturgeon, many cyprinids (bream, carp), catfish, gobies, etc. Due to the predominance of small mollusks in their food, some fish are called "mollusk-eaters", such as the Caspian vobla. Areas where, along with other benthic animals (polychaete worms, brittle stars, etc.), mass development of small bivalve mollusks is observed, serve as feeding grounds for various demersal commercial fish.
Mollusks are readily eaten by many large decapod crayfish (lobsters, hermit crabs, crabs), starfish are the original enemies of bivalve mollusks. Commercial oyster banks are periodically cleaned of starfish with the help of special mops, which are dragged along the bottom by small vessels.
An important role is played by bivalves in the diet of commercial Kamchatka "crabs" (Paralithodes kamtschatica).
What are the main structural features of bivalves? To make it easier to understand their structure, imagine a bound book placed spine up. Both halves of the binding will correspond to the right and left shell valves, covering the body of the mollusk from the sides. The spine of the book will be analogous to an elastic external ligament (ligament) connecting both valves on the dorsal side of the shell and at the same time stretching them. The first and last pages of the book correspond to the two lobes of the mantle, covering the body on the right and left sides, and the next two leaves of the book in front and behind will be analogous to two pairs of gills on each side of the body. And finally, between both pairs of gills, the body itself and the leg are located inside - usually a rather large muscular ax-shaped or wedge-shaped organ directed forward; in attached or inactive forms, the leg can turn into a small outgrowth, and, conversely, in actively moving species (for example, cockles), the leg becomes strong, slightly articulated, adapted for moving in soft sandy soil.
The arrangement of the body parts of a bivalve mollusk will become clearer if we consider an open mollusk, such as toothless, common in our freshwater reservoirs with a muddy bottom and slowly flowing or stagnant water. The most common is common toothless(Anodonta cygnea) is a rather large mollusk from detachment of true laminabranchs(Eulamellibranchia). When examining a mollusk, it is important to determine the anterior and posterior ends of the shell. The front end of the toothless is easily recognizable by the more rounded shape of the shell and by the forward-facing leg; at the posterior, somewhat narrower end, short outgrowths of the mantle-siphons are visible between the valves. Along the upper dorsal edge, behind the tops, there is a rather large external ligament, or ligament, - an elastic elastic cord, with the "reduction" of which the valves open. It consists of a fibrous horny substance close to chitin - conchiolin: it is formed from the outer cover of the shell itself (periostraca). The "work" of the ligament is determined by the interaction of differently located conchiolin fibers, of which it is composed. When the closing muscles, contracting, tighten the shell valves, the fibers in the lower part of the ligament are compressed, and in the upper part they are stretched, and when the muscles are relaxed, vice versa; therefore, in dead molluscs, the shell valves are always half open. In bivalve molluscs, the ligament may be external or internal, or both.
The anodont has no lock teeth and the dorsal edge is smooth, hence its name, toothless (Anodonta). In most bivalve mollusks, in order to more firmly connect the valves to each other under the crown, from the inside, on the dorsal or hinge edge of the shell, there are various (in shape, number and location) outgrowths, the so-called teeth, each of which enters into a corresponding recess on the opposite sash. All this, taken together, forms a shell lock. The device of the lock, the nature, number and location of its teeth is an important systematic feature in bivalve mollusks and is characteristic of the family, genus and species. The ligament is also part of the locking apparatus of bivalves, since it serves to connect the valves to each other.
The shell surface of most bivalve molluscs, including the toothless one, is covered with a differently colored outer layer, or periostraca. It is easily scraped off with a knife, and then a white porcelain-like, or prismatic, calcareous layer (ostracum) is exposed under it. Concentric lines are clearly visible on it - traces of growth of the shell, running parallel to its edges. The inner surface of the shell of many molluscs, including the toothless, is lined with a mother-of-pearl layer (hypostracum).
Periostrak, consisting of conchiolin, is resistant to external influences (both mechanical and chemical) and thus serves as a good protection for the inner calcareous layer of the shell. The resistance of periostraca to the action of carbonic acid dissolved in sea water is especially noticeable. It can accumulate near the bottom, in the very bottom layers, and in the soil where mollusks live (due to the decomposition of organic matter, partly due to the respiration of aquatic organisms), and increase with increasing depth and pressure. So, in the Kara Sea, very soft dead shells of astarte, joldium or portlandium are often found with a dissolved calcareous part of the shell, from which only one intact soft horny layer, the periostracum, remained.
Both other layers of the shell consist of calcareous prismlets or plates connected by a small amount of conchiolin. In the middle (porcelain) layer, they are located perpendicular to the surface of the shell, and in the inner (mother-of-pearl) layer, they are parallel to it; thanks to this arrangement, the interference of light is obtained, which gives the shine and iridescent play of mother-of-pearl. The thinner the plates of this layer, the more beautiful and brighter this shine. The most beautiful mother-of-pearl occurs in those mollusks in which the thickness of the mother-of-pearl plates in the layer is 0.4-0.6 microns.
The shell of a mollusk is formed as a result of the secretory work of its mantle: along its edge there are a large number of glandular cells that produce various layers of the shell. So, the cells of a special mantle groove, running along the entire edge of the mantle, form the outer conchiolin layer, epithelial cells the so-called marginal fold gives the prismatic layer of the shell, and the outer surface of the mantle highlights the mother-of-pearl layer.
The shell of bivalve molluscs, consisting of more than 90% of CaCO3, contains it in the form of calcite or aragonite, which are in various proportions. In tropical mollusks, the shell contains more aragonite, and also quite a lot of strontium. Crystallographic study of the composition of shells of fossil mollusks now makes it possible to judge the temperature of the seas in which these mollusks lived.
Calcium, which is deposited in the shell by the mantle, enters it not only through the blood, where it enters from food through the intestines, but, as recent experiments with radioactive calcium have shown, mantle cells can themselves extract calcium from water.
The growth of the shell occurs both by a general thickening of the valves as a result of the layering of more and more new calcareous plates on the inner surface of the valves, and by the growth of the entire shell along its free edge. When unfavorable conditions occur (in winter, when nutrition deteriorates, etc.), shell growth slows down or even stops, which is clearly visible on the surface of the shell in many mollusks, where characteristic thickening of lines is formed at this time, having the form of concentric stripes, running parallel to the ventral margin of the shell. By winter rings - seasonal growth stops - it is sometimes possible to determine the approximate age of a mollusk. However, in some species such rings are indistinguishable; in tropical forms, where there are no seasonal phenomena, such rings usually do not form at all. Our freshwater barley and toothless have such seasonal winter breaks in growth, so the annual rings are usually well expressed.
To open the valves of the shell of a toothless, it is necessary first of all to cut the two rather strong closing muscles that are inside it in front and behind, which tighten both valves in the transverse direction and close the shell. In a living toothless, it is easier to break its thin shell than to open it without cutting these muscles.
When the muscles are cut, the valves themselves open freely, stretched by the ligament, and one can see two soft translucent pinkish or yellowish lobes - a mantle covering the body from the sides. The edges of the mantle are slightly thickened. In this place, it is attached to the shell, on the inner surface of the valves of which the so-called mantle line is formed. The mantle of the toothless fuses behind to form two short siphons pubescent with short sensitive outgrowths.
Mollusks that burrow into the ground form long contractile siphons; the places of attachment of the muscles that draw them in form an imprint on the inner surface of the shell, the so-called sinus. The deeper the sinus, the longer the siphons of the mollusks, the deeper they can dig into the ground.
On the ventral side, a rather large wedge-shaped leg protrudes forward from under the edge of the mantle, pointing forward with its sharp end. The toothless leg is very mobile (like many other mollusks), and its action is easy to observe in the aquarium. As soon as the anodont has calmed down, its shell opens slightly, the pinkish-yellow edges of the mantle are shown, and the tip of the leg protrudes outward. If everything around is calm, the leg protrudes even further (for large anodonts by 4-5 cm), sinks into the sand, and the mollusk begins to move forward or dig into the ground with its front end, slightly rising on its leg. On the path it has traveled, a trace remains in the form of a shallow groove.
The great mobility of the toothless leg is mainly due to the contraction of the various groups of smooth muscles present in it. There are paired anterior and posterior muscles: retractors that pull the leg obliquely upward, protractors that push the leg forward, and a group of smaller muscles-lifters (elevators) of the leg upward. All these muscles are attached to the inner surface of the shell valves, where the imprints of their attachment sites are quite clearly visible (near the retractors along the hinge edge of the shell). In addition, there are a number of smaller muscles in the leg that are not attached to the valves and are located in the leg in layers and transversely.

If you turn the mantle lobe up, then the mantle cavity of the toothless will open, where its main organs are located: oral lobes, brownish gill sheets (two on each side of the body), a leg, the base of which is located between the right and left gills. In front, in the depression between the leg and the anterior muscle, the mouth opening is placed, surrounded by two pairs of small triangular contractible perioral lobes. Each gill of an anodont consists of two sheets of semigills, which in turn are composed of two plates - ascending and descending.

Each gill plate consists of rows of individual threads (filaments), and each thread, respectively, forms an ascending and descending knee. Anodonts have vascular connections (bridges) between adjacent filaments and between the knees formed by them, which is characteristic of the entire order of true lamellar gills. Each semi-gill is thus a trellis, complexly perforated two-layer plate.
In representatives of other orders of bivalve mollusks, the gills have a different arrangement (which will be discussed below).
The mantle cavity and the gills located in it are constantly washed by the current of water, which is created mainly by the flickering of the smallest cilia of the epithelium covering the surface of the mantle, gills, oral lobes and body walls. Water enters the mantle cavity of the toothless through the lower (respiratory) siphon, first entering its large, lower part - the respiratory chamber, then it is filtered through the gaps in the gills and goes into upper part mantle cavity - into the exhalation chamber, from where it finally exits through the upper (output, or anal) siphon. Water is sucked in through the inlet siphon as a result of the difference in hydrostatic pressure between the subgill and supragill spaces of the mantle cavity and between it and the water surrounding the mollusk; this difference is caused not only by the work of the ciliary epithelium, but also by the contraction of the gill filaments and the muscles of the mantle and siphons. When the flow of water slows down, entering the large "inhalation chamber" of the mantle, coarse and large particles fall out of it and settle on the surface of the mantle, and then are removed from the mollusk. It is easy to verify the presence of intense currents of water entering the mantle cavity, if you put the toothless in a shallow vessel with water so that the water only slightly covers the shell. After letting it calm down, it is necessary to pour into the water near its rear end some kind of powder that remains suspended in the water (for example, ink, carmine, dry grated algae). Then you can see how the powder grains go through the lower (inlet) siphon into the shell and after a while with a strong jet of water are thrown out through the upper (output) siphon. From time to time, toothless, often without any external irritation, slams the shell valves with force and throws out jets of water, renewing all the water contained in the mantle cavity at once. Soon the shell flaps open again and the normal slow circulation of water resumes.
To verify the intensity of the work of the ciliary epithelium, you can cut out a piece of the toothless mantle and put it with the inner surface down on the bottom of the vessel. Due to the work of the cilia continuing for some time, this piece will move slightly and even crawl slightly along the inclined plane.

The suction of water and its circulation inside the mantle cavity provide the toothless not only with the oxygen necessary for its breathing, but also with food. Like all bivalve molluscs, the toothless one is devoid of a head and a number of organs associated with it - a separate pharynx, salivary glands, hard formations for chewing food (such as chitinous plates - a grater found in gastropods). Therefore, toothless can not eat large organisms. She and most bivalves (Eulamellibranchia and Filibranchia) are active filter feeders. Such mollusks feed on detritus suspended in the water column (the smallest remains of dead plants and animals) and microplankton (unicellular algae, bacteria and very small animals). With the help of a complex ciliary mechanism of gills and near-mouth lobes, mollusks filter them out of the water, separating inedible mineral suspension and large food particles for them.

The gill filaments of mollusks have rows of cilia of different sizes located in certain places, which can filter and sort food particles, envelop them with mucus, and then direct them to food grooves located along the ventral edge of the semi-gills (at the transition points of descending gill knees to ascending) or at their base. Rows of rather large lateral, most intensively working cilia on the gill filaments filter water through narrow gaps between the gill filaments and ensure its passage from the "inhalation" to the "exhalation" chamber of the mantle cavity. Particularly large lateral-frontal cilia, located on the sides of the gill filaments, strain food particles from the water or catch them in abundantly excreted mucus and push them to the outer side of the gill filaments. Here are the frontal cilia, which collect food particles and direct them down to the food groove. Food particles that collect in the food grooves are also enveloped in mucus, form lumps here, compact and, thanks to the work of the cilia of the groove, are directed to the oral lobes. The mouth lobes of mollusks are a very efficient sorting apparatus, freeing food from inedible particles. They are armed with many sensitive elements - chemo and mechanoreceptors. They have rows of transverse grooves, armed with especially long cilia; the smallest particles suitable for nutrition are guided along a series of such grooves to the oral groove (located at the base of both lobes), along which they are further directed to the oral opening, where they are swallowed. Along other grooves (with cilia working in the opposite direction to that of the previous ones), larger particles and slimy lumps, unsuitable for nutrition, roll down and fall onto the mantle. The strong cilia of the mantle margins drive these particles back to the base of the introductory siphon; as they move there, these particles stick together, compact and are thrown out in the form of so-called pseudofeces.
In bivalve molluscs from the Protobranchia group (walnut, yoldium, portlandium, etc.), which have the most simply arranged petal-shaped gills - ten and di and, the mouth tentacles are very large, contractile and equipped with a long grooved outgrowth. With the help of it, they collect small food particles from the soil surface - detritus, which are then transferred by cilia along the groove to the plates of the oral tentacles, where they are sorted; gills-ctenidia serve mainly to create currents of water. The work of the filtering and sorting apparatus of bivalves is quite perfect. So, mussels can filter out particles ranging in size from 40 to 1.5-2 microns (best of all - 7-8 microns), completely removing them from the water. They detain unicellular algae and flagellates; heavier particles of mineral suspensions, even 4-5 microns in size, are not retained by mussels. From a mixture of algae and purple bacteria, oysters extract only algae; they usually trap flagellates, algae and organic particles larger than 2-3 microns and let all particles 1 micron or smaller pass through.
Bivalves filter very large volumes of water. So, an oyster can filter about 10 liters of water in an hour; mussel - up to 2-5 liters (at a higher temperature there is more water, at a lower temperature - less); edible cockle at a water temperature of 17-19.5 ° C - from 0.2 to 2.5 liters, an average of 0.5 liters of water per hour; small scallops filter at a rate of 1 liter per hour per 1 g of their weight, while old ones filter only 0.7 liters.
The digestive system of the toothless, like that of all bivalves, consists of a short esophagus, a more or less rounded stomach, middle and hindgut; the ducts of a paired digestive gland, the liver, open into the stomach, and the end of the so-called crystalline stalk protrudes on the ventral side. The intestine (midgut) departing from the stomach at the base of the leg, forms 1-2 turns in the mass of the gonads, then passes to the dorsal side and, penetrating the lower wall of the pericardial sac, passes through the ventricle of the heart, goes beyond the pericardium through its dorsal part, passes above the posterior closing muscle and ends with an anus opening into the cloacal chamber of the mantle cavity with its excretory siphon. The part of the intestine that runs from the pericardium to the anus is commonly called the rectum or hindgut. The intestinal tract of bivalve mollusks does not have muscle fibers, and the movement of food in it occurs due to the work of the ciliary epithelium lining it. The removal of undigested residues is facilitated by the muscular ligament surrounding the anus.
Once through the short esophagus into the stomach, food particles are sorted into small and large due to the activity of the ciliary current and the gastric groove. Large food particles enter the intestines, while smaller ones are carried along the folds of the stomach and are collected at the protruding end of the crystalline stalk. Its protruding end rotates all the time, which contributes to the mixing of food particles and their sorting. The crystalline stalk is formed in a special sac-like organ and is a vitreous rod of a gelatinous substance consisting of a globulin-type protein with enzymes (amylase, etc.) adsorbed in it, capable of digesting carbohydrates (starch, glycogen). Once in the slightly acidic environment of the intestine, it begins to dissolve and release the enzymes adsorbed in it - the only ones secreted by bivalve mollusks in intestinal tract for extracellular digestion of food. Small food particles, processed by enzymes of the crystalline stalk, come from the stomach to the outgrowths of the liver. It consists of a very large number of elongated blind tubules - a diverticulum and is not a digestive gland in the usual sense; it does not produce or release any digestive enzymes into the intestinal tract, and is an organ for intracellular (rather than extracellular) digestion and absorption. Intracellular digestion in bivalves is carried out mainly by phagocytic wandering cells - amoebocytes. They are found in abundance not only in the diverticula of the liver, but also in the stomach and midgut. Amebocytes have various enzymes and are able to digest not only carbohydrates, but also proteins and fats, etc. Wandering cells can pass through the epithelium of the intestinal tract into its lumen and return back to the tissues. Liver cells also swallow and digest food particles; they can also wander through the lumen of the diverticulum and return back to the liver walls. Wandering cells play the main role in the digestion of food in bivalves.
With the death of amoebocytes and liver cells, their digestive enzymes can enter the lumen of the intestinal tract. Therefore, traces of various enzymes (proteases, lipases) are found in extracts from the liver and stomach of bivalves.
Not all organisms that enter the intestinal tract are digested by bivalves. Often, especially with a large amount of food, live diatoms (unicellular algae with a silicon skeleton), small copepods, etc. are found in the fecal masses of mollusks. the concentration of planktonic algae they feed on.
From what has been said, it can be seen that digestion in bivalve mollusks is very peculiar. They can only digest carbohydrates extracellularly, and the protein and fat components of their food are digested by phagocytic wandering amoebocytes and their "liver" cells. Thus, bivalves are a very specialized group of animals adapted to feeding on detritus, unicellular algae, and bacteria.
The circulatory system in toothless, like in all bivalve molluscs, is open, and blood - hemolymph - circulates not only through the blood vessels - arteries and veins, but also in the spaces between organs, and in the connective tissue through a whole system of lacunae and sinuses that do not have their own walls. Arterial blood flows mainly through the vessels, and the venous system has a predominantly lacunar character. Blood is driven through the entire system by the contraction of the heart as well as the musculature of the body. The heart of bivalves (anodonts) consists of a ventricle and two atria and lies in the pericardial cavity, or pericardial sac, located on the dorsal side of the body. The pericardium is an elongated thin-walled sac filled with hemolymph, and in bivalve mollusks it is part of their secondary body cavity, which is greatly reduced in volume. The ventricle has powerful muscular walls and looks like a pear-shaped bag, with its wide end facing backwards. The atria are very thin-walled, translucent and most often look like elongated triangles, the tops of which open into the ventricle; at the entrance to the latter, they are equipped with small semilunar folds - valves that allow blood to pass only from the atrium to the ventricle.
In toothless, like in most bivalves, the ventricle is pierced by the posterior intestine passing through it, but its cavity is completely closed and separated from the intestine by its wall. From the ventricle, blood diverges throughout the body: to the posterior end - through the posterior aorta, which is divided into two arteries that feed the vessels of the posterior part of the mantle and the posterior closing muscle; to the anterior end - through the anterior aorta and arteries extending from it to the leg, to the viscera and to the front of the mantle. From the arterial vessels, the blood pours into the gaps not filled with tissues, and through the system of lacunae, the blood that has become venous is collected through the sinuses and veins into a large longitudinal venous sinus, which lies between the excretory organs. Passing from here through the venous system of the kidneys, it merges into the afferent paired branchial arteries, passing at the base of each gill. Of these, venous blood flows through the afferent gill vessels of the descending gill plates along the gill filaments and their vascular lintels. Arterial blood oxidized in the gills, saturated with oxygen, flows through the efferent vessels of the ascending gill plates into the paired (on each side of the mollusk) gill veins, from where it enters the atria. The atria also receive that part of the blood that, bypassing the gills and kidneys, was oxidized in the vessels of the mantle folds and entered the external gill veins through the mantle veins. In bivalve mollusks, the mantle, with its highly branched blood vessels, plays a very important role in respiration and oxygenation of the blood.
The fact that in most bivalve mollusks the ventricle of the heart is permeated with the rectum finds its explanation in the peculiarities of their embryonic development and in the evolution of this entire group. A number of lower representatives of the bivalves have not only two atria, but also two separate ventricles lying on the sides of the intestine (near the arches); in others, the unpaired ventricle lies above the intestine (nutlets, anomia, limae), in others, it lies down from the intestine (oysters, pearl oysters, etc.) - All this indicates that the location of the intestine and heart in relation to each other has undergone great changes in the evolution of bivalves, and the fact that they originally had two separate ventricles, which then merged together. The frequency of heart contraction in bivalves, which are generally sedentary organisms, is small, usually no more than 15-30 times per minute, while in such mobile and active mollusks as cephalopods, the heart contracts 40-80 times per minute. All parts of the heart of bivalves can contract autonomously.
In bivalves, as in invertebrates with an open circulatory system in general, blood pressure is very low and highly variable.
The blood-hemolymph of bivalves plays a huge role in their life and metabolism. It performs a number of functions: it provides internal organs and tissues with oxygen and nutrients, carries away their metabolic products (carbon dioxide, nitrogen metabolism products, etc.), creates and maintains the constancy of their internal environment (ionic composition, osmotic pressure). Finally, it plays a very important role in creating the hydraulic mechanism of pressure, the necessary turgor (tension), as well as in the movement of mollusks. The study of blood circulation in the body of bivalve mollusks explained the phenomenon of swelling of the legs, which is observed when the animal moves and burrows. It occurs due to its filling with blood, which gives the leg the necessary elasticity, creates the necessary turgor. When the leg is stretched and the leg muscles relax, blood flows through the artery to the leg, and when it contracts, it goes back to the body. So, in a sea stalk, which can burrow very quickly, the leg first sinks into the ground and the blood quickly flows into it, expanding the end of the leg in the form of a disk; the latter serves as an anchor when the leg muscles, contracting, pull the mollusk down. When the mollusk rises from the ground to the surface, the leg muscles relax, and the end of the leg expands again, filling with blood; holding on to such an "anchor", the leg is extended, as part of the blood enters the upper part of the leg and pushes the mollusk up. The inflow, injection of the amount of blood necessary for swelling of the leg and its outflow are regulated by the so-called keberian organ, which plays the role of a valve.
In contrast to animals with a closed circulatory system, bivalve mollusks, like all invertebrates with an open circulatory system, have a fairly significant amount of blood - hemolymph. In mollusks (except cephalopods), it is 40-60% (volume percent) of their body weight without a shell. At freshwater pearl mussel(Margaritifera) and mussels(Mytilus californianus) per 100 g of body weight, the volume of blood is about 50 ml.
In the blood of bivalve mollusks there are many formed elements, mainly various forms of amoebocytes (leukocytes). Their number varies in different species from 6,000 to 40,000 per 1 mm3. Bivalves also have erythrocytes; sometimes they can even be more than some forms of leukocytes. Hemoglobin is found in quite a few species (arches, sea cuttings, tellins, pectunculus, astartes, etc.).
Important for gas exchange (for supplying organs and tissues with oxygen), the ability of blood to be saturated with oxygen in bivalve mollusks is very small and amounts to 1-5% of their blood volume. So, 100 ml of toothless blood can absorb only 0.7 ml of oxygen, while in mussels - 0.3 ml. Toothless consumes 0.002 ml 02 per 1 g of its weight in one hour (at 10°C); oysters, respectively - 0.006 ml 02 (at 20°C); mussels-0.055 ml 02. More mobile species usually consume somewhat more, such as, for example, scallops Pecten grandis, consuming 0.07 ml 02 per 1 g of their weight in 1 hour (or 70 cm3 02 per 1 kg of weight). Small forms also often consume more oxygen than large ones. For example, at the optimum water temperature (18°C), the horny charr consumes 0.05 mg 02 per 1 g of weight per hour, but when the water temperature drops to 0.5°C, oxygen consumption almost stops. In oxygen consumption, i.e., in the intensity of metabolism, many bivalves show seasonal fluctuations; Thus, mussels in the summer, the most active time of their life, consume approximately twice as much oxygen as in the winter, cold season.
Many bivalves can live for quite a long time with very little or no oxygen in the water. So, sand shell(Mua arenaria) can live in anoxic conditions at 14°C for up to 8 days, and at 0° even for several weeks; virgin oyster also endures such conditions for a week or more. Metabolism during such periods of anaerobiosis is sharply reduced, but molluscs can at the same time receive the oxygen necessary for their life through intramolecular respiration - glycolytic breakdown of their reserve substances (carbohydrates and fats) by the type of fermentation. This ability for temporary (facultative) anaerobiosis (anoxibiosis) is especially characteristic and necessary for species living in the littoral zone (such as, for example, for sand shells, mussels, Baltic poppy, edible cockle). At low tide, they close their shells, a small amount of oxygen in their mantle cavity disappears rather quickly, and they begin to live due to the processes of anoxybiosis. At high tide, they open their shells, continuously filter the water and breathe in the oxygen dissolved in the water; at first, they sharply (several times) increase the intensity of filtration and oxygen consumption, and then after a while it returns to normal, characteristic of their life in water.
The organs of excretion in bivalves are the kidneys, and also, but to a lesser extent, the so-called keber organ; the latter is a glandular thickening of the anterior part and anterior-lateral walls of the pericardial sac.
The kidneys, or Boyanus organs, open with their inner ends into the pericardium, and with their outer ends into the mantle cavity. Toothless kidneys look like two dark green curved tubular sacs; one end has glandular walls and represents the actual excretory part of the kidney, the other has the form of a bubble, where metabolic products accumulate to be removed from the body.
In bivalves, there is no such concentration of parts of the central nervous system (nerve nodules, or ganglia) as in gastropods. The toothless, for example, has one pair of ganglia above the mouth, a little behind it, another pair deep in the leg, and a third behind the posterior locking muscle. Between the first and second pair of ganglia, as well as between the first and third, a pair of nerve trunks passes, and each pair of nodules is interconnected by transverse bridges (commissures).
The sense organs in bivalves are rather poorly developed compared to other classes of mollusks. However, these organs are quite diverse in their structure and are scattered in different parts of the body: along the outer edges of the mantle, at the ends of the siphons, on the first gill filaments, near the mouth opening on the near-mouth tentacles, on the edges of the posterior closing muscle, in the exhalation chamber, near the back intestines, etc. These sense organs are both rather complex formations - eyes, or photoreceptors, balance organs - statocysts, or statoreceptors, and simpler ones - osphradia, various sensitive outgrowths, and sometimes simply clusters of pigmented sensitive cells.
Photoreceptors in bivalves can be arranged very differently: from simple epithelial pigment (optical organelles) to rather complex eyes with a lens and retina. Such eyes can be very numerous, especially in free-living forms, such as the mantle eyes in combs, in which there are sometimes up to 100 of them on both edges of the mantle.
Differently arranged eyes and ocelli can also be located in bivalves on the first gill filaments (gill eyes in arches, anomies), on short outgrowths around siphon openings (in some cockles, etc.).
Many bivalves have so-called optical organelles, spherical or elongated, concentrating light on a special intracellular innervated formation (retinella). Such photoreceptors are scattered at the ends of siphons and in other parts of the body of molluscs.
The balance organs in bivalves are a vesicular protrusion of the epithelium, well innervated, lined from the inside with ciliated epithelium, closed (statocyst) or open (statocrypt). They contain hard mineral grains (statolith) or small grains of sand (statoconia) inside. Usually, for example, in toothless, statocysts are located near the foot ganglion or on the dorsal side of the mollusk. Balance organs are well developed in free-living forms, such as scallops.
Osphradia are usually very small paired pigmented, well innervated groups of sensitive cells. They can be located in various places - on the leg, in the region of the gills, hindgut, etc. Their role is still not clear enough: whether they are chemoreceptors or organs of touch.

Anodonts, like most bivalves, have separate sexes, but in the conditions of reservoirs with stagnant water, individual hermaphroditic individuals or even their entire colonies can be found. At the same time, in order to avoid self-fertilization, male reproductive products are first produced - spermatozoa, and then female - eggs. Paired, strongly dissected sex glands (gonads) in bivalves (including anodonts) lie in the dorsal part of the leg, where they are surrounded by an intestinal loop and outgrowths of the liver; excretory ducts gonads open into the mantle cavity near the openings of the excretory system. Only in some of the most primitive bivalves, the gonadal ducts open with a common opening with an excretory pore. In some freshwater bivalves, sexual dimorphism is so pronounced that males and females of the same species are sometimes referred to as different species.

The development of juveniles in bivalve mollusks is varied. Almost all marine forms that live in shallow waters lay their eggs directly into the water, where fertilization occurs, or it occurs in the mantle cavity of the mother. Eggs float freely in water, rarely stick together or attach to shells, algae. The exceptions are viviparous (more precisely, larval-bearing) forms, like some oysters, arches, etc.
Fertilized eggs of bivalve mollusks, having passed the stage of crushing of the spiral type, form a trochophore-like larva, similar to the larva of polychaete worms (polychaetes). However, in the process of embryonic development of bivalves, there is almost no segmentation process, which is so characteristic of the development of the larval stages of annelids. The larvae of bivalve molluscs have an rudiment of a leg and a primary shell (prodissoconch), which is initially laid down as a single plate located on the dorsal side of the larva. After a series of changes in the trochophore, in which the sail (velum) arises - the ciliary parietal disc, the bivalve shell and the rudiments of other organs, turn into a veliger. The presence of a free-swimming larva (veliger) is a very important life stage of mollusks, as it provides them with the possibility of wide dispersal, since adult bivalves usually lead a sedentary or even attached lifestyle. At the same time, this larval stage of life, like that of other invertebrates, is the most sensitive to unfavorable external conditions, and only the high fecundity of bivalve mollusks ensures the preservation of the species and its distribution.
In a number of marine cold-water and, apparently, in many deep-sea oceanic species of bivalve molluscs, development can occur with a very shortened stage of the floating larva or without it at all. In the latter case, a few large eggs are formed, supplied with a large amount of nutrients. This allows them to develop regardless of the presence of food in the surrounding water, which is especially important for deep-sea forms, where the amount of food for juveniles at the bottom is very limited.
Based on the structure of the shell, castle, gills, on the location and number of muscle-contacts, ligaments, etc., detachments are distinguished: Comb-toothed, Ligamentous-toothed, True lamellar-branched and Septoid-branched.
Animal life: in 6 volumes. - M.: Enlightenment. Edited by professors N.A. Gladkov, A.V. Mikheev. 1970 .
Bivalve mollusks (Bivalvia) are distinguished by a shell consisting of two parts (flaps). This class has several more names that characterize the representatives of this class. For example, Lamellibranchia are lamella-gill molluscs whose gills are actually made of plates. Acephala are headless mollusks that have lost their heads in the process of evolution. Pelecypoda (axepoda) - the name describes the shape of the limbs of bivalves.
 There are several types of bivalve molluscs that have a name that describes their external features.
There are several types of bivalve molluscs that have a name that describes their external features. Lifestyle features
The second largest group of mollusks after gastropods consists of more than 20 thousand species. All these species are benthic, i.e. bottom. Bivalve mollusks live at the bottom of reservoirs with fresh or salt water. Most Bivalvia are extremely slow or lead an almost immobile life.
For example, the speed of movement along the bottom of representatives of the species of river bivalve mollusks - toothless - is no more than 20-30 cm per hour. And oysters, for example, are attached to the substrate even in the larval stage and are unable to move at all.
Evolutionary changes associated with the disappearance of the head and radula (from the Latin radula - scraper, scraper, grater for scraping food) and the formation of developed lamellar gills led to the establishment of such a little or no motionless lifestyle.
Bivalve mollusks lead an inactive lifestyle at the bottom of water bodies.
The depths at which different species of the Bivalvia class live vary from the tidal coastal zone to sea trenches 10 km deep.
Bivalve molluscs feed on organic particles and small plankton. Filtering water suspension with the help of gills, they implement two functions at once: respiration, absorbing oxygen from the water, and nutrition, filtering out edible particles.
Some groups of laminabranchs have interesting adaptations to life on rocks. Species belonging to the genus Pholas, for drilling passages in stones, have sharp teeth at the front end of the shell. And another species of marine bivalve mollusks, called the sea date (Lithophaga), although it does not have a drilling apparatus, is also able to penetrate stones, dissolving them with acid, which is secreted by special glands.
Body structure and shell
The body of a mollusk is placed inside a bivalve shell. consisting of a body and a leg. The leg is a muscular part of the body, with the help of which the molluscs move along the bottom or burrow into the ground. It often has a wedge-shaped shape and is able to protrude from the shell.
Inside the shell is the body of the mollusk
Many species, such as the mussel (Mytilus), have a byssal gland in the leg that secretes a substance that helps the mollusk attach itself to stones and similar substrates. Byssus is a strong thread. Some adult mollusks do not have such a gland, in which case it was most likely developed at the larval stage.
Shells of laminabranchs can have various sizes and shapes. The smallest deep-sea mollusks do not grow more than 0.5 mm in length. But there are also giants, for example, tridacna - an inhabitant of coral reefs in tropical seas. The size of this type of bivalve can reach 1.4 m in length with a body weight of up to 200 kg.
Most species have an oblong body flattened laterally. But there are also species with an elongated worm-like or almost spherical shape. The sink can be symmetrical or have valves of different sizes. Most representatives of bivalves have a lock on the shell valves, which prevents the valves from moving relative to each other.
Regardless of the shape and size, the shell consists of three layers:
- external - conchiolin;
- internal - lime;
- lower - mother-of-pearl.
The sea shell of a bivalve mollusk is thicker than that of a freshwater inhabitant
The thickness and strength of the shell in different species is also different and depends on habitat conditions. A large amount of minerals in the water allows you to build up a more durable calcareous skeleton, so marine bivalves usually have a thicker shell than freshwater species. The part of the body of the mollusk adjacent to the valves secretes substances that make up the shell. Thus, over the course of life, the shell gradually grows. Among Bivalvia with a well-developed mother-of-pearl layer, there are freshwater species (barley, freshwater pearl oyster, etc.) and marine species (sea pearl oyster, etc.).
The practical importance of bivalves
People living in close proximity to the shores of seas and rivers have long used Bivalvia as food. And from their shells and pearls they made household utensils and jewelry. Many laminabranchs are used for consumption as food. The most common types are:
- mussels (Mytilus);
- cockles (Cardium);
- oysters (Ostrea);
- scallops (Pecten).
Pearl fishing
When a foreign irritant enters the shell, a pearl is formed
At present, the mariculture of Bivalvia mollusks is widely developed, i.e. their artificial breeding. They are grown for the purpose of using them for food or for obtaining pearls.
Founded in 1907 in Japan, the company was the first production of cultured pearls. Bivalvia was mined for this in the open sea, and only in the mid-50s was it possible to establish the cultivation of pearl mussels themselves.
Foreign objects placed in the shell of a pearl oyster are gradually enveloped in mother-of-pearl. And after 1-2 years, it is possible to extract finished pearls, which are carefully sorted by size and shade and sent to enterprises for the manufacture of jewelry.
Biological water treatment
The ability of Bivalvia mollusks to biofilter is considered a useful property of these living organisms. The direction considering the possibility of using these animals for water purification is considered relevant. Mollusks are able to absorb and accumulate heavy metals in their body tissues and purify water from chemical and organic contaminants. The average activity of lamellar gills during water filtration is about 1 liter per hour.
One of the most useful properties of these organisms is the ability to purify water
The issue of protection and breeding of Bivalvia for use as biofilters in fresh and marine waters is considered by scientists as one of the the most pressing issues. In areas where commercial breeding of laminabranchs is established, high-quality biological purification of water takes place, bottom silt accumulates, the richest benthic fauna develops, and the overall productivity of the ocean increases.
Sedimentary rock formation
Dying mollusks form calcareous sedimentary rocks, which form layers on the sea and ocean floor, which are the material for the formation of shell rock, marble, limestone. Shell fossils are the forms upon which the determination of the age of the earth's layers is based.
Malicious Representatives
First of all, Bivalvia mollusks harm hydraulic structures and sea ships. Active development of special coatings capable of protecting ships and structures from pest fouling is underway.
Some species of mollusks are pests
In the rivers and sea waters of the Black and Caspian Seas, where the bivalve mollusk of the species Dreissena Polymorpha lives, significant colonies of these animals can form, which are attached to hydraulic structures. These animals settle in water pipes and turbines of hydroelectric power plants, leading to blockages.
A well-known pest is the mollusk Bivalvia of the species Teredo navalis (or cones), also called the shipworm. It is found in the Black and Far Eastern seas, reaches a length of 18 cm and has a worm-like shape. The sink occupies only one end and is adapted for drilling wood. The mollusk damages wooden structures and the bottoms of ships. To combat the woodworm, the tree is tarred.
The number of species is about 20 thousand. Habitat - sea and fresh waters.
Consider the features of the class using the example of its representative - toothless. The general scheme of the structure of the toothless is shown in fig. 9.61.
Rice. 9.61.
1 - the line along which the mantle is cut; 2 - muscle-closing; 3 - mouth; 4 - leg; 5 - oral lobes; 6,7 - gills; 8 - mantle; 9 - inlet siphon; 10 - outlet siphon; 11 - hindgut; 12 - pericardium
The body is completely covered by a shell consisting of two valves. The shell has anterior (blunt) and posterior (pointed) ends, dorsal and ventral margins. The shell valves are connected by the dorsal edges with the help of an elastic ligament (it contributes to the opening of the shell, since there are no opening muscles).
The body is located mainly in the dorsal part of the shell, covered with a mantle (the folds of which form siphons - see below).
There is a wedge-shaped leg that serves to move and burrow into the ground.
The head is missing.
The nervous system has a simplified structure: three pairs of ganglions connected by commissures, and nerves extending from them.
The sense organs are poorly developed, represented by primitive balance organs and chemical sense organs - sensitive cells on the gills, in the wall of the mantle and siphons. Some representatives of the class have photoreceptor cells along the edges of the mantle.
The digestive system is based on the nutrition of protozoa, unicellular algae, suspended in water. Food with a stream of water through the introductory siphon enters the mantle cavity, where it is filtered from mineral particles, after which it is transported to the mouth, then to the stomach (into which the ducts of the bilobed liver flow), the middle and hindgut, ending with the anus, which opens into the mantle cavity. Excrement from the latter is removed with a stream of water through the outlet siphon.
Due to the reduction of the head of the pharynx, tongue and salivary glands are absent.
The circulatory system is of an open type.
The heart is three-chambered (two atria and one ventricle).
Respiratory system: gills, consisting of thin plates, braided with a dense network of blood capillaries.
The excretory system is an unpaired kidney.
There is a division.
Sex glands are paired.
Fertilization is external (spermatozoa from the mantle cavity of males are brought out through the siphon, and then with a stream of water enter the mantle cavity of the female, where fertilization occurs).
Other representatives of the class: barley, mussel, Far Eastern scallop, shipworm, zebra mussel, oyster, tridacna (the largest representative of the class: length - 140 cm, weight - 250 kg).
The role of bivalves in nature and human life:
- are a link in food chains, water purifiers;
- accelerate the transfer of C, P, N through aquatic ecosystems, filtering out significant amounts of suspensions containing these elements and depositing them in the form of biological nodules on the bottom of water bodies;
- are pests of agricultural crops (slugs, grape snails);
- cause damage (by damaging wood) to ships and port facilities (shipworm);
- used as a food product (mussels, oysters, scallops);
- used to make jewelry from mother-of-pearl and pearls.
Class Cephalopods
The number of species is about 700.
Habitat: seas, oceans.
The scheme of the body structure of cephalopods is shown in fig. 9.62.
Rice. 9.62. The structure of a two-gill cephalopod mollusk (using the example of a female cuttlefish):

1 - ink bag; 2 - part of the body cavity; 3 - pericardial cavity; 4 - heart; 5 - kidney; 6 - gill; 7 - the opening of the kidney into the pericardial cavity; 8 - external opening of the kidney; 9 - genital opening; 10 - anus; 11 - mantle; 12 - funnel; 13 - more dexterous tentacle; 14 - tentacle; 15 - pharynx with jaws; 16 - ganglia; 17 - eye; 18 - esophagus; 19 - salivary gland; 20 - liver; 21 - stomach; 22 - shell rudiment; 23 - ovary; 24 - beginning of the oviduct
There is a head and a body.
The leg is turned into tentacles that have shifted onto the head and surround the mouth opening, or a muscular tube - a funnel (siphon).
There is a shell: a) in primitive forms - external multi-chamber (nautilus pompilus); b) in higher forms - internal, reduced (octopuses).
The body is surrounded by a mantle.
The integument consists of skin (single-layer cylindrical epithelium) and dermis with chromatophores (color change occurs by changing the shape of these cells).
Propulsion system: 10 tentacles with suction cups (2 tentacles - trapping, 8 - for capturing prey and other manipulations).
The muscles of the mantle and the funnel serve to push water through the funnel (siphon) (the consequence of this is a reactive effect that provides a high speed of movement).
The nervous system is a large brain (the result of the fusion of the ganglia of the peripharyngeal ring), enclosed in a cartilaginous capsule.
Sense organs:
- two large eyes (derivatives of the integument; the structure is similar to the human eye, accommodation is carried out by moving the lens relative to the retina);
- olfactory pits (under the eyes);
- organs of balance (inside the cartilaginous skull);
- taste organs (chemoreceptors on the tentacles).
The structure of the digestive system is associated with heme, that cephalopods are carnivores (predators).
The scheme of functioning of the digestive system is as follows:
mouth (2 horny jaws, tongue with a grater, salivary glands with a poisonous secret) -» pharynx -> esophagus -? stomach, where the liver ducts open small intestine -? hindgut (the ink gland opens here) -? powder (opens into the mantle cavity).
Respiratory system: gills (2-4) inside the mantle cavity. The circulatory system is almost closed (there are few gaps, they are short).
In the blood - the pigment hemocyanin, the composition of the molecules includes Si, which gives the blood a blue color;
Structure circulatory system shown in fig. 9.63.

Rice. 9.63.
Excretory system: 2 or 4 kidneys.
The reproductive system and reproduction are characterized by the following.
There is a division.
Sex glands are unpaired.
In females, the genital duct opens into the mantle cavity, where fertilization occurs.
In males, gametes enter a special spermatophore bag, in which spermatozoa stick together and form special packages - spermatophores. Spermatophores are introduced into the mantle cavity of the female with the help of a special tentacle (hectocotyl).
Eggs are laid in special nests.
The development is direct.
Class members:
- subclass Fourgills(ancient and primitive). Example: nautilus (the heart consists of 1 ventricle, 4 atria), there is a multi-chamber shell;
- subclass Bibranchial(the most organized). Examples: octopuses, squids, cuttlefish, argonauts (the heart has 1 ventricle, 2 atria).